- КУЛЬТУРЫ ТКАНЕЙ
- КУЛЬТУРЫ ТКАНЕЙ. Содержание: История..................... 74 Методика и техника.............. 75 Общая биология тканевых культур...... 82 Применение метода тканевых культур в различных областях биологии и медицины . . 90 Культуры тканей (син.: эксплян-тация тканей, тканевые культуры), выращивание тканей вне организма, in vitro. Под именем метода тканевых культур подразумевают методику и технику, сущность к-рых состоит в следующем: небольшой кусочек ткани или органа, свеже вырезанный из организма, помещают в подходящую среду при таких условиях, чтобы не только сохранить жизнь ткани (простое «переживание»), но и получить настоящий рост ткани (размножение клеток) вне организма. Такой метод экспериментирования в сочетании с разнообразными способами изучения получаемых таким путем «тканевых культур» оказался весьма плодотворным и нашел широкое применение в разных областях цитологии, гистологии, эмбриологии, биохимии, физиологии, патологии, фармакологии, бактериологии, иммунологии, гематологии, радиологии и т. д. Подобно тому как пересадку части одного организма на другой (или на другое место того же организма) обозначают термином «трансплян-тация», так и помещение части организма в какую-либо другую среду, но не в живой организм, называют (согласно термину, предложенному Roux и введенному в употребление Оппелем) эксплянтацией. Следовательно тканевые культуры представляют собой такие тканевые эксплянтаты, к-рые в отличие от разных других эксплянтатов обнаружи- вают в том или ином виде рост тканей. Хотя возможна эксплянтация и целого зародыша (Totalexplantation), целого сердца зародыша и т. п., но при экспериментах с тканевыми культурами имеют дело обыкновенно с эксплянтацией небольших частей (Teilex-plantat, Teilzuchtung). Для изолирования и выращивания тканей могут' быть использованы и другие методы: можно по способу Экмана, Штёра, Бауцмана и др. поместить кусочек зародыша, зачаток сердца и т. п. в мешочек, вырезанный из эктодермы (обыкновенно личинки амфибии Bombinator); при этом внутри эктодермального мешочка растут и развиваются различные эмбриональные ткани. Изолирование и выращивание in vivo эмбриональных тканей («тканевые культуры in vivo») удается при пересадке кусочка зародыша на хорио-аллантоис куриного зародыша, развивающегося в яйце (Murphy, Данчакова и др.), при имплянта-ции в брюшную полость, в полость глазницы и т. п. (см. Иитерплянтация). В дальнейшем изложены данные только о тканевых культурах in vitro в указанном выше смысле. История. Хотя отдельные попытки выращивания тканей вне организма предпринимались в разное время многими авторами (Roux, Борн, Л. Леб, Габерландт и др.), однако основателем современной методики тканевых культур следует признать Р. Гар-рисона (R. Harrison). Он первый правильно. установил основные принципы выращивания тканей вне организма (необходимость опоры для движения и роста клеток, малую величину эксплянтированных тканей для облегчения обмена веществ,'необходимость стерильности среды и асептики всех манипуляций и т. п.) и впервые (1907) опубликовал удачные опыты выращивания тканей лягушечьего зародыша в капле свернувшейся лимфы лягушки. Берроуз (Burrows), работая в лаборатории Гаррисона, улучшил метод: он начал применять вместо лимфы плазму крови. В последующих работах Карреля и Берроуза (1910), производившихся уже в Рокфеллеровском ин-те в Нью Иорке, описаны тканевые культуры из различных тканей и органов разных животных, а также человека. Чтобы продлить рост тканевых культур, к-рый в первоначальных культурах продолжался лишь 3—15 дней, Каррель и Берроуз начали производить «пассажи», т. е. промывали эксплянтированные кусочки ткани и помещали их затем в новую среду. Особенное значение для дальнейшего развития успехов в этой области имело открытие Каррелем (1913) того факта, что эмбриональный сок обладает способностью усиливать рост тканей. Применение в качестве среды плазмы крови с прибавлением эмбрионального сока (что и в наст, время признается наилучшей средой) и систематическое проведение «пассажей» дали возможность Каррелю и другим установить существенно важный биол. факт, что соматические клетки разных животных и человека способны расти вне организма неограниченно долгое время. Т. о. современная методика и техника могут быть названы «методом Гар-рисона-Берроуза-Карреля» (менее правильно называть их «методом Карреля»). При помощи разработанного Каррелем метода пассажей, которые производятся каждые 2—3 дня (см. ниже), до наст, времени поддерживается рост тканевых культур, которые были приготовлены в Рокфеллеровском ин-те 17/1 1912 года из сердца 7-дневного куриного зародыша. В течение первых пятнадцати лет с этими культурами эмбриональных фибробла-стов было проделано около 3.000 пассажей. В настоящее время эти культуры, живущие вне организма 18 лет, обнаруживают прежний хороший рост без малейших признаков старения и могут считаться «вечными».-—Альберту Фишеру (Fischer) в 1921 году впервые удалось получить чистую культуру эпителия из радужной оболочки глаза куриного зародыша; затем были получены чистые штаммы эпителия щитовидной железы, хондриобластов, моноцитов и некоторых других клеток. Особенно важным успехом было получение чистых штаммов злокачественных опухолей: куриной саркомы (Альбертом Фишером в 1924 году), чистой культуры раковых клеток из перевиваемой Эрлиховской мышиной аденокарцпномы (им же в 1927 г.), саркоматозных клеток в чистой культуре из 2 крысиных сарком (Каррелем в 1927 г.). Возможность экспериментировать с чистыми штаммами тканей и производить количественный учет результатов опытов посредством измерения скорости роста (Каррель, Ebeling) или путем анализа обмена веществ (Кронтовский и его сотрудники) сделала выращивание ткани in vitro более точным методом научного исследования и способствовала более широкому и плодотворному применению метода таковых культур в различных областях. Усовершенствование методики и техники происходило и в других направлениях: для исследований физиол. характера Каррель (1923) предложил новый метод, по к-рому кусочек ткани эксплянтируется в двухслойную среду (см. ниже) таким образом, что рост ткани непрерывно продолжается в плотном слое среды, а верхний жидкий слой среды периодически возобновляется. Максимов (1916) тщательно разработал гистолог, технику исследования тканевых культур; Кронтовским (1925) введены физ.-хим. и микрохим. методы для изучения жизнедеятельности, хим. динамики тканевых культур, в том числе и не растущих (напр. эксплянтатов из разных частей головного мозга взрослых животных и т. д.). Опыты с культурами из тканей и органов разных животных и человека в последнее время так широко и настолько часто я систематически применяются для изучения различных вопросов биологии и медицины, что в 1925 г. при участии специалистов из разных стран был основан специальный интернациональный журнал «Arch. i. exper. Zellforschung besonders G-evve-beziichtung», а в сентябре 1927 года во время X Международного конгресса зоологов в Будапеште была создана особая секция, посвященная работам в этой области. В разных государствах имеются специально оборудованные лаборатории, приспособленные для работ с чистыми штаммами тканей (с пассажными культурами): лаборатории А. Карреля в Рокфеллеровском институте в Ныо Иорке, отделение А. Фишера при Биол. ин-те в Берлин-Дамлее, лаборатории (завед. А. А. Кронтовский) в Бакт. ин-те и Рентген, и радиолог. ин-те в Киеве, гистол. лаборатории Дж. Леви в Анатомическом ин-те в Турине. В последнее время за границей и в СССР аналогичные лаборатории основываются при различных ин-тах. Методика и техника. Оборудование специальных лабораторий и общая установка работ с тканевыми культурами осуществляются или 1) согласно принципам хир. асептики (комната, аналогичная хир. операционной, стерильные халаты, шапки, маски на лице у экспериментатора, на столе в качестве фона большой стерильный кусок черной материи и т. п.)г и тогда нек-рые манипуляции, например вырезывание из тканевой культуры кусочков для последующего пас- сажа, производятся без всякой защиты от загрязнений через воздух и т. п., или же* 2) соответственно принципам экспериментальной бактериологии, когда приготовление тканевых культур, пассажи и т. д. проводят в обыкн. лабораторной комнате, достаточно чистой (экспериментатор может работать без стерильного халата, маски и пр.), но все манипуляции производят при строгом соблюдении правил бактериол. чистоты: культуры вырезываются под защитой (от загрязнений воздушным путем) крышки от чашки Петри, слегка приподнятой с одной стороны для введения инструментов; жидкости берутся согласно бактериол. правилам и т. п. При достаточном навыке хорошие результаты получаются при организации работы и по первому и по второму типу. При всех работах с тканевыми культурами вся посуда, растворы, инструменты и т. п. должны быть простерилизованы, а все манипуляции производиться асептически. 1. Основные типы приготовления тканевых культур. Хотя тканевые культуры могут быть приготовлены в различной посуде, разными способами, однако в последнее время выкристаллизовалось два основных типа приготовления тканевых культур, к-рые и следует применять (по возможности всегда в одинаковых «стандартизованных» условиях, если по сути дела для данного случая не требуется особой техники): 1) тканевые культуры (или вообще эксплянтаты) по типу висячей капли (см. рисунок в т. V, ст. 89); 2) тканевые культуры в чашке Карреля в двухслойной среде [см. рисунок 1 и отд. табл. (ст. 79—80), рис. 1 и 2]. Можно эти типы комбинировать. Для р ^ ,^г\ продления жизни тка- r-*t----------^^-^шШ) невой культуры неко- У^^=~^\^^ Bf^f торые авторы предло- ^^^^РИ^5""^ ^ ^ жили спец. аппара- ё А ТЫ С непрерыв- Рисунок 1. Схема ткане-Н О Й Ц И р К у Л Я Ц И - вой культуры в чаш-ей, В КОТОРЫХ расту- ке Карреля. щая ткань постоянно омывается протекающей жидкостью: каким-нибудь физиологическим раствором, сывороткой, жидкостью эксудата и т. п. (аппараты Берроуза, Ро-мейса, Васильева, де Гаана и др.); однако при помощи метода пассажей (см. ниже) или сменой жидкой части среды в чашках Карреля продление жизни достигается более успешно, в виду чего названные выше аппараты и не получили практич. применения. Для приготовления ткапевых культур по первому типу лучше всего постоянно пользоваться специальными толстыми предметными стеклами с углублением одинакового размера (для облегчения сравнения результатов с данными разных авторов и т. п.), а именно: 26x76 мм с круглым углублением определенной глубины (б. ч. 3—4 мм) с диаметром в 20— 21 мм и покровными стеклами или пластинками слюды (к-рые лучше переносят стерилизацию) 24x40 мм.— В последнее время Каррель предложил готовить одновременно 4 тканевых культуры по типу висячей капли накруглой (5 см в диаметре) пластинке слюды, к-рая потом укрепляется в качестве крышки на металлическом круге (ободе), причем дно такой чашки также представляет собой пластинку слюды (см. ниже). Можно покровные стекла с культурами помещать вчашки Петри, готовить эксплянтаты в чашках Габричевского и т. д.—Для культур 2-го типа применяются как правило чашки Карреля (см.отд.табл., рис. 1 и 2) с одним горлышком, с диаметром чашки в 3 си (тип D.,) или в 5 см (D6). Плотная часть среды (рис. 1—а), в которой находится экплянтируемый кусочек ткани (рис. 1—е), состоит из плазмы крови с примесью разведенного эмбрионального сока; жидкая часть среды (рисунок 1—Ъ)—обыкновенно из жидкости Тироде •.(Tyrode) с 5—10% эмбрионального сока (или из гепариновой плазмы, иногда из сыворотки), Для получения очень медленного ро-■ста (что способствует развитию некоторых гистол. структур) Фишер применяет иногда новую модификацию—прибавляет в чашку Карреля ничтожное количество разведенного эмбрионального сока, нужного лишь для -лучшего свертывания плазмы, а жидкую В часть среды после промывания совсем отсасывает. Кроме основных типов тканевых культур предложено также множество модификаций: выращива-. ние тканей в пробирках по способу Карреля, Риверса, Гаагена и Муккенфуса (Rivers, Haagen u. ^^ Muckenfuss) [рисунок 2 (а и Ъ—два Рисунок 2. слоя среды; с — тканевая культура)]. по Шампи (Champy), в флаконе Борреля (Borrel), в специальных ка-жерах, сосудах и т. д. 2. Среды для тканевых культур. Наилучшей средой для тканевых культур до настоящ. времени считается свернувшаяся плазма крови (в к-рой сеть фибрина является хорошей опорой для передвигающихся и размножающихся клеток), смешанная обыкновенно с эмбриональным соком (или,с разведенным эмбриональным экстрактом), иногда с протеозами и т. п. Плазма крови получается следующим образом. Берется кровь посредством промасленной канюли у животного (чаще всего у курицы) из art. carotis или при помощи шприца (смазанного внутри вазелином) из сердца маленьких животных или из вены руки человека. Кровь набирается в парафинированные центрифужные пробирки (или под слой жидкого парафина), стоящие на льду. Кровь центрифугируется в течение нескольких минут на электрической центрифуге (можно пользоваться я ручной). Прозрачная плазма, собравшаяся над осевшими эритроцитами и лейкоцитами, снимается и идет сейчас же для приготовления культур или сохраняется в запасе во .льду. Если работают с «гепариновой плазмой» (см. Гепарин), то можно обойтись без парафинирования и охлаждения. Обыкновенно применяют плазму животного того же вида, что и культивируемая ткань. Можно однако пользоваться и гетерологическими плазмами, успешно выращивать ткани аксолотля, например в плазме кролика (Кронтовскии и Шустова, Хлопин), или ткани мыши—в плазме курицы (Фишер) л т. п. В общем плазма курицы по заключению Карреля является наилучшей средой для выращивания тканей животных разных видов, в том числе и млекопитающих (включая и человека). Т. к. можно применять и гетерологические эмбриональные экстракты <Каррель), то для культивирования тканей разных животных и человека особенно часто пользуются ■смесью плазмы курицы с куриным эмбриональным •соком. Цопидимому последний может быть заменен раствором «протеоз» или продуктами (типа пептона Witte) непродолжительного пепсинного переварива--ния фибрина или же тканей разных органов (см. ниже). Нередко в качестве среды пользуются и одной плазмой, обыкновенно разведенной, раствором Тироде •или Рингер-Локка или же с прибавлением тканевых -экстрактов из костного Мозга (Максимов), из селезенки (Кронтовскии), из опухолей (Drew), из дрожжей <Heaton). Необходимо иметь в виду, что фибробласты и нормальный эпителий безгранично могут расти <с размножением клеток) лишь при наличии в среде эмбрионального сока (хотя бы разведенного), тогда как моноциты и ткани нек-рых опухолей могут раюти и без него (напр. в плазме с сывороткой). В известных •случаях можно пользоваться комбинированными средами по Кронтовскому. Если по заданиям исследования желательно иметь тканевые культуры в жидких средах (т. к. плазма нередко сильно затрудняет специальные исследования, напр. при темнопольном освещении, при разных микроскопическо-химических пробах и т. п.), то пользуются средой Локк-Лыоиса, одним эмбриональным экстрактом, гемолимфой (Гольдшмидт), жидкостью Дру (см. Дру раствор), смесью сыворотки с раствором Тироде и т. п.; но для получения роста жидкая среда должна быть распределена очень тонким слоем, чтобы ткань и клетки не плавали в капле жидкости, а были прижаты к стеклу, к-рое и будет служить опорой; в указанных жидких средах растут обычно лищь эмбриональные ткани (от зародышей ранних стадиев, напр. куриные зародыши от 5 до 11 дней) и лишь недолгое время. Прибавление фибриногена (Ebeling) оказалось непрактичным; вместо этого Каррель в' последнее время пользуется свернувшейся плазмой (в чашке Карреля), из к-рой сыворотка удалена промыванием. Эмбриональный с'ок каждый раз приготовляют свежий: чаще всего берут 7—10-дневный куриный зародыш, измельчают его на кусочки и центрифугируют; полученный эмбриональный сок обыкновенно разводят раствором Тироде или Рингер-Локка. При приготовлении сред обращают внимание па то, чтобы среда была изотонична (или слегка гипотонична), имела бы соответствующую реакцию, и т. д. 3. Приготовление тканевых культур. Ткани для эксплянтации препарируются при соблюдении строгой асептики в растворе Тироде или Рингер-Локка острыми инструментами (катарактальным— глазным—ножом Грефе или небольшими ножницами) и разрезаются на маленькие кусочки с поперечником чаще всего приблизительно в 1 мм (0,5—2,0 мм). В зависимости от разных условий размеры эксплянтата могут варьировать. Для эксплянтации лейкоцитов крови поступают обыкновенно так: нормальную или лейкемическую кровь человека или животных сильно центрифугируют, плазму отсасывают, на оставшийся слой лейкоцитов (расположенный над эритроцитами) спускают несколько капель эмбрионального сока; когда лейкоцитарный слой свернется в виде пленки, ее промывают и режут на кусочки и эксплянтируют, как и кусочки ткани. Можно воспользоваться для эксплянтации и клетками эксудатов. Сравнительно легко удаются тканевые культуры из перевиваемых опухолей животных. Если опухоль разжижает плазму, то предпочтительнее культивировать ее по методу Фишера, помещая в среду рядом с кусочками опухоли кусочек мышцы (или какого-нибудь органа). При эксплянтации тканей растений материал получают или механическим путем, вырезая маленькие кусочки ткани, или же изолированием при помощи плазмолиза. Средой служит чаще всего раствор Кнопа с добавлением глюкозы, «пептона Witte» и т. п., к к-рому прибавляется иногда и немного агара: в такой среде Котте, Шустова и др. наблюдали рост кусочков корешков гороха, выращенных стерильными при помощи особых приемов. Многие авторы (в особенности Габерландт и его ученики) эксплянгировали кусочки картофельных клубней, . кусочки листьев и т. д. 4. Метод пассажей, разработанный Каррелем, применяется для поддержания длительной (безграничной) жизни штаммов тканей, особенно—в чистой культуре. Пассаж производится таким образом, что у хорошо разросшейся (обыкновенно 48-часовой тканевой культуры) (рис. 3—а) четырьмя прямыми разрезами отрезаются периферические части новой выросшей зоны, а оставшаяся средняя часть (прежний кусочек с каемкой новой ткани) (рис. 3—Ь) промывается в жидкости Тироде, и из нее готовится новая тканевая культура (субкультура), к-рая в течение 48-час. роста опять разрастается до прежней величины (рис. 3—с); затем снова производят пассаж и т. д. Можно хорошо разросшуюся культуру разрезать на 2 части (рис. 4—1) и затем половину каждого кусочка (рис. 4—2) можно эксплян- l^jUJ т l»;v•,, ' {\ ,*
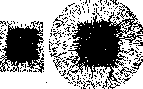
Рисунок 3. Пассажные тканевые культуры.
тировать в виде отдельной культуры (рис. 4—3); такие культуры обнаруживают обыкновенно почти одинаковый рост (рисунок 4, кривые роста), в виду чего они являются очень точным биол. реагентом для изучения всевозможных воздействий, к-рым подвергается одна из них (другая оставляется для контроля). Иногда удается вырезать часть новообразованной зоны и из нее получить дальнейший рост. Пассажи делают обычно каждые два-три дня; при работе с тканями холоднокровных, растущими при комнатной темп. —значительно реже, например каждые 2-—2Va недели. Пассажную тканевую культуру, к-рая росла в течение 24 ч. в термостате, можно потом на много дней (до 25 дней по опытам Мейе-ра) оставить при комнатной t° или на холоду, а затем с успехом сделать пассаж. Это обстоятельство да-возможность перевозить или пересылать почте такие культуры на очень далекое / / *ш, ) / ?Н0 / м* - / t ' г # Я4" у/ ', ='U часы Рисунок 4. Рост двух тканевых культур, приготовленных из одной культуры фибробла-стов. ет ПО расстояние. 5. Исследование тканевых культур, а) Наблюдение живых культур. Особо важное значение имеет микроскоп, исследование живых тканевых культур: кратковременный осмотр производится просто при комнатной t°; для длительного наблюдения за амебоидной подвижностью,.делением клеток, ритмическим сокращением тканевых культур из сердца и т. п. эксплянтаты ставят на согревательный столик, или микроскоп вместе с культурой помещается в специальном термостатном ящике. Важную услугу оказывает исследование при темнопольном освещении, т. к. при нем в живой клетке видны митохондрии (хондриосомы), заметно появление различных изменений в структуре цитоплазмы и ядра и т. д. [см. отд. табл. (т. XIV, ст. 375— 376), рисунки 6 и 7]. Для получения отчетливых картин (при темнопольном освещении) необходимо специальным образом монтировать препарат или применять Preparier-Wechsel-Condensor'bi Петерфи (фирмы Цейс). Прекрасные картины получаются при помощи методов прижизненного окрашивания. Зарисовывание и микрофотографирование производятся в общем обычным образом. Для целей микрокинематографии, к-рая позволяет проследить тончайшие детали движения нормальных и раковых клеток, их размножения и т. д., в последнее время предложена сравнительно простая аппаратура. Тканевые культуры особенно пригодны и для различных микроопераций на клетках, для микрургии (см.), т. к. при этом клетки во время операции находятся, так сказать, в более «физиол.» условиях, чем. при других методах, и после операции можно проследить клетку в условиях тканевых культур в течение долгого времени; для регистрации процесса микрооперации и последствий ее особенно пригоден фотографич. окуляр (например «Phoku» Zeiss). Для проведения микрургии (см.) в тканевых культурах необходимы некоторые специальные приемы и приспособления, разработанные Петерфи, Кронтовским и др. б) Фиксирование и окраска тканевых культур для цитологических и г и ст. целей. Для получения микроскопических (тотальных) препаратов из целых тканевых культур [к-рые для получения хороших препаратов должны быть заранее (при изготовлении тканевых культур) приготовлены достаточно тонкими, не содержать толстого слоя плазмы или приготовлены в жидких средах] Максимов рекомендует фиксацию культур в жидкости Корнуа (для тонких препаратов—жидкость Ценкера) и окраску гематоксилином Делафильда. Каррель и его ученики пользуются обыкновенно формалином. Фишер фиксирует тканевые культуры в 2%-ном формалине (в Рингеровском растворе) или в 2%-ном формалиновом алкоголе, окрашивает гематоксилином, промывает, а затем через спирт, ацетон-ксилол, ксилол доводит до канадского бальзама. Для получения окрашенных срезов Максимов применяет фиксацию тканевых культур почти исключительно в Zenker-Formol'е (15—20 мин.) и быстро заключает в целлоидин. Леви для фиксации тканевых культур особенно рекомендует жидкость Максимова такого состава: к 8 см3 основного (запасного) раствора Ценкера прибавить 1 см* чистого формалина и 1 ш' 2%-ной осмиевой к-ты. Фиксировать 2.—6 мин. После фиксации—заключение в целлоидин или целлоидин-парафин по Петерфи (в парафине не более 15 минут) и окраска Гейденгайновским железным гематоксилином. На основании тщательных исследований Стрепд-жуейса и Канти (см. ниже), посвященных изучению изменения живых клеток при фиксации, Нивен (Niven) предварительно подвергает тканевые культуры действию 2%-ного раствора осмиевой кислоты, затем фиксирует в жидкости Ценкера (без уксусной к-ты) и окрашивает железным гематоксилином. Для специальных целей применяются соответствующие методы фиксации и окраски. в) Биол. и физиол. методы. При исследовании жизнедеятельности тканевых культур и применении их для изучения различных вопросов биологии и медицины особенно важное значение имеют количественные методы. Жизнедеятельность тканевых культур можно измерять различным образом: интенсивностью обмена веществ, скоростью роста, количеством митозов и т. п. Так как увеличение массы ткани растущей культуры измерить непосредственно не удается (в виду трудности взвешивания), то о ней судят по увеличению поверхности тканевой культуры,, причем скорость роста выражается обычно по методу Иблинга: зарисовывают (при помощи проекционного рисовального аппарата) контуры только что эксплянтированного кусочка (рис. 4—2) и определяют при помощи планиметра поверхность (А0) после 48 часов роста (или вообще за время t), опять зарисовывают контуры всей разросшейся куль- 6 - 7 Рисунок 1 и 2. Чашка Карреля с тканевой культурой. Рисунок 3. Чистая культура эмбриональных фиброблае-тов (край зоны роста). Рисунок 4. Эпителиальные клетки из чистой культуры эпителия (iris куриного зародыша). Рисунок 5. Эксплянтат кусочка 10-дневного зародыша кролика с «органотипическим» ростом (внизу-обыкновенное врастание ткани в среду). Рисунок 6. Чистая культура ракового эпителия (из аденокарци-номы мыши). Рисунок 7. Смешанная культура, содержащая эпителий и соединительную ткань, полученная in vitro из чистых штаммов эпителия и фибробластов. К ст. Культура тканей. туры (рис. 4—3) и определяют всю площадь (At), т. ч. новообразованная зона будет равняться At—А0. Тогда индекс Иблинга («относительное увеличение» тканевой культу- At — А ры) =г ". При изучении различных воз- ■"■ о действий отношение показателя роста в данном эксперименте (Е) к показателю роста контрольной культуры (К) количественно выражает влияние изучаемых воздействий на рост. Вместо «относительного увеличения» по Иблингу, по вычислениям Бух-Андерсена и Фишера правильнее пользоваться VА или log VA, где А есть поверхность выросшей культуры. Для суждения об интенсивности обмена определяют или потребление определенных веществ—сахара, кислорода и др. или образование известных продуктов—молочной к-ты, увеличение кислотности среды, изменение рН (Кронтовский). Особенно удобным показателем интенсивности обмена в эксплянтатах из разных тканей, растущих и нерастущих (напр. из разных частей головного мозга), показателем изменения жизнедеятельности под влиянием лучей радия (Кронтовский), изменений в содержании К и Са (Яцимирская-Кронтов-ская) и т. д. оказалось потребление сахара (предпочтительнее—вместе с определением образования молочной кислоты). Для определения жизнедеятельности экеплянта-тов по методу Кронтовского к среде прибавляются вещества (напр. глюкоза и другие углеводы, трибути-рин и т. п.), к-рые под влиянием жизнедеятельности тканей способны разлагаться с образованием продуктов, вызывающих изменение реакции среды, и по истечении известного срока измеряют изменение рН. Для изучения влияния разных тканей на рост («бла-стотропных» влияний), на полярность роста и т. п. Чентанни (Ccntanni) применяет метод противопоставленных («аффронтированных») эксплянтатов: в одной капле среды на расстоянии приблизительно 1 мм помещают 2 кусочка одинаковых или разных тканей. Другой способ Чентанни: на таком же расстоянии от экспляитированного кусочка укрепляют маленький стеклянный капиляр, наполненный испытуемой жидкостью. Из физиологических методов можно отметить способ Кронтовского регистрации на ленте кимографа сокращений эксплянтатов из сердца, движения мерцательного эпителия, стандартизацию фармакол. веществ по остановке сокращений сердечных эксплянтатов, перистальтических движений кусочков кишки (Фишер) и т. п. В общем современная техника предоставляет экспериментатору самые разнообразные возможности, позволяет готовить тканевые культуры из различных тканей животных и человека, получать очень интенсивный равномерный рост (напр. в пассажных тканевых культурах или в чашке Карреля) или же культуры с очень медленным ростом (по модификации Фишера), получать разрастание сравнительно простой однородной («чистой») ткани (ст. н. цитотипическим или гистотипическим ростом) или же рост и развитие сложных, органоподобных или даже организмоподобных эксплянтатов (см. ниже) и исследовать тканевые культуры различными морфол. и физиол. методами. При работе с методом тканевых культур требуются критическое отношение к производимым исследованиям и осторожность в выводах, т.к. одни явления протекают как в тканевых культурах, так и в организме в общем одинаково (проявления прижизненной окраски, поведение клеток соединительной ткани и крови и их дальнейшее превращение и т. п.), другие—иначе (напр. реализация эффекта действия лучей Рентгена в организме по сравнению с тканевыми культурами, не всегда одинаковая обратимость повреждений, нанесенных клеткам в организме—при тран-сплянтации и вне организма—в эксплянтатах и т. д.). И для надлежащей постановки опытов и для получения правильных выводов нужны соответствующие знания об особенностях тканевых культур, правильная оценка преимуществ и недостатков техники, знание границ применения метода. В соответствующих случаях желательно вводить параллельно и другие методы исследования. Общая биология тканевых культур. — 1. Общая морфология. Если экс-плянтирован кусочек куриного зародыша, то уже через несколько часов по краям кусочка в окружающую среду в большом количестве начинают врастать (б. ч. радиаль-но) отдельные соединительнотканные веретенообразные или отростчатые клетки («соединительнотканный тип роста»), т. ч. уже через 48 часов вокруг кусочка образуется обыкновенно широкая (см. рисунок 3—а и с и отд. табл., рис. 3) новообразованная зона, т.н. «зона роста» (или «инвазионная зона»— зона внедрения). Эпителий растет обычно в виде сплошного тонкого и широкого пласта, мембраны («эпителиальный тип роста»—-см. отд. табл., рис. 6), состоящей из эпителиальных клеток (см. отдельную таблицу, рис. 4). Эпителий образует иногда тяжи, выстилает стенки полости, образующейся нередко в плазме. По Рубашкину и Шахову, для эпителия характерен в общем рост в виде сим-пласта (сплошного слоя), для соединительной ткани—синцитиальныи рост; и в том и в другом могут обособляться отдельные клетки. Часто в культурах имеет место смешанный рост. Иногда наблюдается вырастание мышечных волокон с многоядерным расширением на конце (как при регенерации) и нервных волокон, периферический конец которых расширен и снабжен тончайшими, нитевидными подвижными амебоидными отростками. При эксплянтации кусочков селезенки, лимфаткч. узлов, костного мозга, лейкоцитарной пленки и т. п. уже через несколько часов из кусочка в окружающую среду эмигрируют различные блуждающие клетки—лимфоциты, лейкоциты, ретикулярные клетки и т. п., так что сначала «зона роста» состоит лишь из эмигрировавших клеток; только позднее из кусочка начинают расти веретенообразные фибро-бласты, и потом наступает действительное размножение клеток, появляются митозы в гипертрофированных ретикулярных клетках (макрофагах), в фибробластах и других клетках. Вообще следует различать более сложные явления, развивающиеся внутри самого экспляитированного кусочка (в случае железы—явления в железистых участках, выводных протоках, разрастание эпителиальных трубок, пролиферация интер-стициальной ткани, процессы прогрессивного и регрессивного характера, доходящие в центре кусочка до некроза и т. п.), и внешнее разрастание (в среде) вокруг кусочка более простого эпителиального пласта (см. отдельн. табл., рис. 4 и 6). В известных слу- чаях (например при культивировании кроличьих зародышей ранних стадиев) обычного роста культур — врастания тканей в окружающую среду—совсем не происходит, а кусочки зародыша округляются, поверхность их обрастает эпителием (см. отд. табл., рисунок 5, верхняя и боковые части), внутренние ткани обнаруживают иногда органо-идное развитие; получается как бы своеобразный растущий организм (похожий нередко на тератому). Такие эксплянтаты Максимов предложил называть «органотипиче-скими» или «организмотипическими» в отличие от обыкновенных, характеризующихся «гистотипическим» или «цитотипическим» ростом. 2. Тканевая к у льтура как биологическая система. Тканевая культура не является простым агрегатом отдельных независимых клеток, а представляет собой своеобразную биол. систему. Отдельные (изолированные) фибробласты напр. и эпителиальн. клетки, судя по опытам Фишера, Поликара и др., не могут размножаться: для их пролиферации необходима известная совокупность, определенное минимальное число клеток [это обстоятельство Фишер ставит в связь с наличием гипотетических дес-монов (см.), Берроуз—с определенной концентрацией гипотетических веществ—«ар-хузии» и «эргузии» и т. д.]. При сращении двух тканевых культур из кусочков сердца с двумя разными ритмами сокращений получается культура с одним общим ритмом (опыты Фишера, Olivo), т. ч. образуется единая система не только в морфол., но и в фнкц. отношении. По вопросу о связи клеток друг с другом в тканевых культурах следует отметить, что многие авторы описали в них синцитиальные образования и указали, что из этих синцитиев при известных условиях легко обособляются, освобождаются отдельные клетки и эмигрируют в среду. Мнения авторов о характере межклеточных связей расходятся: одни полагают, что отростки клеток только плотно пристают (прилипают) друг к другу (Льюис), так что разные гранулы, напр. при своем движении, не переходят из клетки в клетку (de Garis), тогда как по мнению других между клетками существует непосредственное протоплаз-матическое соединение (Фишер). Результаты опытов с повреждением клеток иглой микроманипулятора в одних случаях (напр. эпителия эпидермиса) говорят за непосредственное соединение (Chambers), в других—против этого (Kredel). В тех случаях когда между соседними клетками заметить границы невозможно, индивидуальность клеток в биол. (физиологическом) смысле по мнению Леви все же сохраняется, т. к. каждое ядро имеет обычно свою собственную сферу влияния. В комплексных тканевых культурах, содержащих разные ткани, как и при сращивании чистых культур разных тканей (например эпителия и соединительной ткани и т. п.), влияние ткани друг на друга (межтканевые корреляции) выражено достаточно ясно. В органотипич. эксплянтатах наблюдается ряд явлений, имеющих характер регуляции (организации): Максимов описал отграничение поверхности эмбриональных эксплянтатов от окружающей среды эпителиальным покровом (см. отд. табл., рис. 5), преобразование тканей, элиминацию эпителием дегенерированных клеток с последующ. восстановлением изъяна и т. д. Округленные, эпителиализированные, замкнутые в себе эксплянтаты Смирнова наблюдала при эксплянтации пальцев аксолотля, Тимофе- евский и Беневоленская—концевых частей конечностей эмбрионов человека, Фишер, Хлопин—кусочков кишки зародыша, слизистых оболочек мочевого пузыря, Умела, Гасуль и др.—кусочков слизистой оболочки зева лягушки, Муррей (Murray) — при эксплянтации кусочков плянарий и т. д. При культивировании лейкоцитов крови разных животных (позвоночных, а также и беспозвоночных) и человека — моноциты (амебоциты и т. п.) и часть лимфоцитов (зернистые лейкоциты скоро погибают) превращаются в макрофагов (см.), в конце-концов даже в фибробластов, причем в культурах лейкоцитов морской свинки Максимов описал образование настоящей соединительной ткани с аргирофильными и коллагенными волокнами (экспериментальный синтез ткани из отдельных клеток). У других (низших) животных из отдельных разъединенных клеток могут получаться не только ткани, но и целый организм: в опытах Гальцова и др. из отдельных искусственно диссоциированных клеток губок (Microciona prolifera и др.) образовались округленные агломераты, к-рые развивались в настоящие маленькие губки, иногда же (при разрушении поверхностной мембраны) становились похожими на обычные тканевые культуры. Путем совместного выращивания чистых штаммов разных тканей получаются биолог, образования (единицы) высшего порядка, напр. наподобие желез (Fischer, Parker и др.) и т. п. (см. отд. табл., рис. 7). В тканевых культурах из зародышей удается наблюдать и развитие нервной симпат. сети, охватывающей целые клеточные области (Максимов), и присоединение выросших нервных волокон к элементам соседнего кусочка мышечной ткани (Лаврентьев, Григорьев). Т. о. метод тканевых культур дает возможность не только расчленять живой организм—получать и анализировать чистые расы клеток, составляющих организм, но и экспериментировать в синтетическом направлении (в смысле экспериментальной «синтезиологии» Гейденгайна). Что касается проблемы диференцировки и деди-ференцировки (обратного развития), то Шампи выдвинул положение, что в тканевых культурах эпителий из разных органов подвергается дедиференциров-ке, превращается в индиферентный эпителий, а потом эпителиальные клетки становятся неотличимыми и от клеток соединительнотканного происхождения. Это положение оспаривается многими авторами (Максимов, Хлопин, Фишер и др.). В наст, время с несомненностью выяснено, что в тканевых культурах из кусочков зародышей, цельных зачатков конечностей, зачатков глаза, слухового пузырька и т. п. наблюдается диференцировка: эктодерма обнаруживает ороговение, образование кожных сосочков, рудиментарных молочных желез и абортивных волосяных фоликулов (Максимов); установлено in vitro образование хряща и кости (Fell), разных частей глаза (Fell, Филатов), функциональная (появление ритмических сокращений), а затем и гист. диференцировка сердечной мышцы (Olivo) и т. д. Обнаружена и биохим. диференцировка: при эксплянтации бедер зародыша, не содержащих фосфатазы, наряду с нормальным развитием in vitro типичных зон хряща, при образовании гипертрофированных клеток появляется в большом количестве и фосфатаза (Fell и Robinson)—фермент, участвующий повидимому в процессе оссификации. Разумеется, зачаток печени, напр. уже в силу пространственных, механических и иных условий, не развивается в настоящую печень—наблюдается лишь разрастание эмбриональных печоночных клеток, кро-'веносных капиляров и т. п. (Максимов, Беневоленская). Установлено также, что чистые штаммы эпителия сохраняют свои основные свойства эпителия в течение неопределенно долгого времени, причем эпителий щитовидной железы напр. может продуцировать коллоид в течение больше 4 месяцев жизни и роста вне организма; эпителий радужки—образовывать пигмент и т. д. С другой стороны в тканевых культурах наблюдается иморфологич. дедиференцировка, известное упрощение строения: эпителиальные клетки простаты со специальными зернышками секрета (Champy) или характерное для разных отделов под-желуд. железы строение железистых клеток (Хлопни) постепенно теряют в тканевых культурах свою органоспецифическую железистую структуру, и эпителиальные пласты, разросшиеся из разных частей железы, дают одинаковый индиферентный эпителий (Хлопин). Дедиференцировка наблюдается и в экс-плянтатах из сердца (Оливо и друг.)- Необходимо однако иметь в виду, что разнообразные изменения (трансформации) формы и характера клеток во многих случаях могут не относиться собственно ни к диференцировке ни к дедиференцировке, а зависят от окружающих условий, от свойств среды (Льюис, Барта и др.), от усиленной пролиферации и т. п. При изменении условий—при сочетании in vitro с другой тканью и т. п.—эпителий, как указано, опять обнаруживает полярную, характерную структуру и т. п. {Максимов, Дру, Фишер и др.). Прибавление соединительной ткани способствует диференцировке эпителия (Дру), невробластов (Лаврентьев, Григорьев). Сращивание культур штамма эмбриональных фибробластов и эпителия, долгое время культивированных в отдельности, причем эпителий вырастал в виде индиферентного эпител. пласта, дало образование наподобие железы (см. отд. табл., рис. 7), с типичным расположением эпителия и соединит, ткани (Фишер и Иблинг). Мало того, получение более диференцированных структур удается и в опытах с чистым штаммом: Фишер и Паркер при интенсивном росте культур, полученных из перихондрия куриного зародыша, наблюдали морфол. дедиференцировку, при искусственно замедленном росте — появление более диференцированной ткани с межклеточным веществом. Очевидно усиленная пролиферация мешает диференцировке, это наблюдается и в организме: при эмбриональном развитии периоды усиленного роста чередуются с периодами диференциров-ки (Шмальгаузен и Степанова).—Условия жизни клеток и тканей в эксплянтатах своеобразны, но во многих отношениях сходны с теми, к-рые имеют место в организме при регенерации, заживлении ран, новообразовании ткани, при воспалении и т. п. (Stran-geways, Максимов). Соответственно этому в эксплянтатах (как и в организме при разных условиях) происходит энергичная мобилизация блуждающих клеток в покое (гистиоцитов), ретикулярных клеток, которые становятся свободными, моноцитов крови, гипертрофия всех этих клеток и превращение их в «макрофаги» («полибласты» Максимова). Самые различные клетки—в том числе пигментные клетки (в эксплянтатах из органов лягушки и аксолотля), хлорагогенные клетки (в культурах из тканей дождевого червя), невробласты (в эмбриональных культурах) и т. п.-—приобретают амебоидную подвижность (она ясно заметна и у края растущего пласта эпителия, у концевого расширения растущего нервного волокна и т, д.) и эмигрируют в среду. Делению подвергаются различные тканевые клетки, лейкоциты нормальной и лейкемической крови; иногда начинают размножаться даже клетки, к-рые в организме обычно не обнаруживают деления, например Мюллеровы клетки сетчатки (Champy). Мясоедов наблюдал в эксплянти- рованных яйцевых фоликулах яичников (кролика) дробление яйца, что позднее описал и Шампи под названием «экспериментальный партеногенез у кролика». 3. Общий тип хим. динамики тканевых культур. Разнообразные клетки и ткани в эксплянтатах (как и в организме при регенерации, заживлении ран, продуктивном пролифер. воспалении и т. п.) обнаруживают (как указано выше) большую активность. Источником необходимой для всего этого энергии служит гл. обр. сахар (глюкоза), как показали исследования Крон-товского и его сотрудников Бронштейна, Яцимирской-Кронтовской, Коломиец и др. (подтвержденные теперь уже и многими другими авторами). Потребление сахара в тканевых культурах так значительно, что маленький кусочек ткани (поверхность которого равна приблизительно 1 мм2), составляющий по весу всего лишь около Veoo—Viooo всей среды, в течение 48 часов поглощает до 60 % (или даже больше) всего содержащегося в среде сахара (рис. 5). . Если содержание сахар б % 0 15 iac. ч ч ч ч ч ч _4Ш>4 iac. ч0.136 \ ч \ \ s \ 0.04а Рисунок 5. Потребление сахара культурами из чистого штамма фибробластов. сахара в среде искусственно (напр. диализом) понижено ниже определенного уровня (0,005% по Винду), то тканевые культуры не обнаруживают роста; прибавление сахара к такой (диализированной) среде делает ее опять пригодной для роста. В самых разнообразных тканевых культурах (и на слюде и в чашках Карреля, даже в атмосфере кислорода) наряду с исчезновением большого количества сахара всегда обнаруживается значительное увеличение количества молочной к-ты. Т. о. хорошо растущая in vitro нормальная ткань покрывает значительную часть своей потребности в энергии за счет брожения (что согласно теории Варбурга присуще исключительно злокачественному росту карцином и сарком). Так как при возбуждении мышц (и в нек-рых других случаях) также выступает сначала анаэробная фаза расщепления углеводов, а возбудимость (раздражимость) представляет собой общее элементарное свойство живой протоплазмы, то Кронтовский высказал теорию, согласно которой при возбуждении разных тканей пускается в ход прежде всего указанный общий энергетический механизм, наблюдается сдвиг в сторону усиления процессов брожения, т. е. процессов, легко дающих энергию, энергия же в зависимости от разных условий, внутрен. структуры и т. д. идет или на сокращение мышцы или на процессы усиленной активности клеток и тканей при процессах регенеративного характера, в тканевых культурах in vitro, при заживлении ран в организме, при воспалении и т. д. При разных опытах с тканевыми культурами для правильного понимания многих явлений нужно иметь в виду, что среда не остается постоянной: уже в течение 2-дневного роста эмбриональных культур содержание сахара резко падает (рис. 5), увеличивается количество молочной к-ты, накопляется С02, происходит сдвиг рН в кислую сторону (в культурах по типу висячей капли рН среды сначала около 8,6, потом в течение-2 дней передвигается до 7,0—6,8 и более в кислую сторону; в чашках Карреля при измерении специальным электродом рН кусочка ткани первоначально = 7,2, затем—7,0— 6,8, а среды—8,1—8,3) и т. д. Накопление СОа происходит отчасти благодаря тканевому дыханию, гл. же обр. вследствие вытеснения С02 из бикарбонатов образовавшейся молочной к-той (что показал Magath прямыми опытами с прибавлением соответствующего количества молочной кислоты в свежеприготовленную, герметически замкнутую культуру). Посредством вентиляции культур в новой (металлической) чашке Карреля с 2 боковыми трубочками (см. выше) или же путем простого временного приподнимания слюды с культурой по типу висячей капли (Магат) можно передвинуть реакцию назад к первоначальному, более щелочному пункту, а вдувание воздуха с примесью 5 % СОа в чашку Карреля может свежую, несколько щелочную среду (рН около 8,0) немного подкислить; таким путем удается до известной степени сделать среду более стабильной в указанном отношении. 4. Анализ роста и питания клеток. Наблюдаемые в тканевых культурах явления—усиленное разложение сахара с образованием молочной кислоты при недостаточности дыхания, вытеснение С02 молочной кислотой из бикарбоната, ведущее при задержке С02 к подкислению,—■ могут служить прототипом, легко доступным изучению, ряда явлений в организме: при ишемии, воспалении, регенерации, заживлении ран, трансплянтации и т. п. и помогают понять их механизм. Культуры чистого штамма «фибробластов» (клеток мезенхимы) и других клеток оказались прекрасным биологич. (физиологическим) реагентом, позволившим открыть и изучить ряд факторов, способствующих росту ткани или задерживающих его. Продолжительный, неограниченный рост фибробластов и эпителия невозможен (по опытам Карреля) в одной плазме или сыворотке; при наличии же в среде эмбрионального сока указанные ткани могут питаться и синтезировать свою протоплазму неограниченно долгое время. Длительное размножение моноцитов, лимфоцитов, клеток нек-рых опухолей возможно и в сыворотке. Способствующие росту вещества содержатся не только в эмбриональном соке, но и в других тканевых экстрактах (из селезенки, из саркомы Rous и т. д.), но только в меньшем количестве. По Каррелю в эмбриональном соке в протеиновой фракции имеются специальные азотсодержащие питательные вещества, которые он назвал «трефонами». Каррель, Бекер (Baker), Фишер, Райт, Гитон (Heaton) и др. тщательно исследовали их природу. Оказалось, что первые стадии («протеозы») пепсинного переваривания (неполного гидролиза) разных протеинов—фибрина, чистого кристаллического альбумина, кристаллического эдестина, разных органов (печени, гипофиза и т. д.) значительно усиливают рост тканей in vitro, иногда не хуже (по Каррелю—даже лучше) эмбрионального экстракта. По мнению Карреля эффект действия эмбрионального сока зависит от того. что в нем легко происходит расщепление клеточными ферментами протеинов сока в протеозы; по Фишеру, эффект действия зависит не от хим. веществ, а скорее от особого физ.-хим. состояния. Если среда составлена из продуктов переваривания чистых веществ (кристаллического альбумина или казеина), то прибавление недостающих в этой смеси гликоколя и нуклеиновой к-ты улучшает рост (Каррель, Бекер, Иблинг). При изучении значения' окси-редукционного потенциала для тканевых культур Бекер обнаружила, что культивирование в синтетической среде (смеси продуктов казеина, гликоколя и нуклеиновой к-ты) дает лучшие результаты при прибавлении к ней глютатиона и гемоглобина (или золы печени). Сыворотка оказывает задерживающее влияние на культуры фибробластов и эпителия и укорачивает их жизнь (Каррель). Дальнейшие исследования привели Карреля к заключению, что в сыворотке имеется некоторое количество и антагонистического принципа, способствующего росту. Задерживающее влияние сыворотки увеличивается с возрастом животного (Каррель и Иблинг). Согласно теории Карреля питание и рост клеток зависят, с одной стороны, от наличия в окружающей их жидкости 2 групп антагонистических субстанций (способствующих росту — «трефонов» — и задерживающих его), с другой стороны — от внутренней энергии данной ткани, к-рая больше у тканей зародышей и молодых животных, меньше—у старых. Представление о внутренней энергии роста ткани может быть получено (но Каррелю) путем измерения т. н. «остаточной энергии роста», т. е. продолжительностью и скоростью роста культуры в среде, не содержащей питательных веществ. По мнению Карреля кривая роста фибробластов и эпителия (рис. 6) в питательной среде, содержащей эмбриональный экстракт, сходна с параболой; в среде, недостаточной для питания, имеет S-образную форму. На основании математического анализа разных, фактически полученных кривых Бух-Андерсен и Фишер считают, что кривых роста первого типа вообще не наблюдалось; всегда замечается большее или меньшее последовательное замедление роста. 1'. к. согласно опытам Карреля лейкоциты in vitro могут вырабатывать из сыворотки вещества, способствующие росту («лейкоцитарные трефоны»), культуры из лейкоцитов способны оказывать на соседние, дегенерирующие культуры фибробластов омолаживающее влияние и т. д., и так как такие же продукты образуются повидимому и в организме (они были обнаружены в экстрактах из воспаленных тканей, в перитонеаль-ном эксудате с макрофагами и т. п.), то Каррель выдвинул гипотезу, согласно к-рой элементы белой крови, особенно же, как это было им выяснено позднее, лимфоциты и моноциты, являются в организме как бы подвижными одноклеточными железами, способными питать фибробласты и эпителий. И вследствие этого именно эти клетки играют большую роль при заживлении ран, при воспалительном разрастании и т. д., так как рост ткани зависит от определенной концентрации в окружающей среде «трефонов», к-рые сецернируются белыми тельцами, а также выделяются из погибших тканей (мышцы и т. д.). Для роста тканей необходимы и многие другие факторы: например определенная реакция среды (для фибробластов optimum рН, по Фишеру, 7,4—7,8), определенное сочетание электролитов [изменение содержания К и Са резко влияет и на рост и на обмен веществ эксплянтированных тканей (Яцимир-ская-Кронтовская)], известное осмотическое давление [3'w ' 9ftO / / £W / / / «0 / n (Hogue), определенная t°, в качестве источника энергии—сахар (глюкоза) и т. д. 5. Основные типы тканевых культур и и х жизненный цикл. По условиям жизни, по их жизненному циклу можно выделить 3 основных типа тканевых культур. 1) Тканевые культуры (экс-плянтаты) с простым элементарным жизненным циклом: маленький кусочек ткани (около 1 мм в поперечнике) помещается в каплю среды (напр. плазмы с эмбриональным соком); в этом случае не производится ни промывания тканевых культур для удаления продуктов обмена ни прибавления новой среды. Тканевые культуры при этих условиях растут (при t° тела) обычно в течение 5—15 дней (в зависимости от различных условий и дольше, ткани холоднокровных напр.—при комнатной t° больше месяца). Затем клетки дегенерируют, и культура гибнет. 2) Тканевые культуры, жизнь которых удлиняется посредством возобновления среды (или при эксплянтации в 2-слойной среде в чашке Карреля путем возобновления ее жидкой части), причем сама растущая часть остается нетронутой (рисунок 1). Рост фибробластов продолжается около 15 дней, причем колонии их достигают больших размеров—2—3 ем в диаметре; рост моноцитов (макрофагов) —■ тридцать и более дней (Кар-рель). Кривая роста эпителия изображена на рис. 6. 3) Тканевые культуры, жизнь которых поддерживается постоянн. пассажами в течение неопределенно долгого времени (пассажные, «вечные» культуры). При пассажах имеет место не только промывание эксплянтата и помещение в новую среду, но кроме того при каждом пассаже отрезаются периферические части (рисунок 3), следовательно наблюдается постоянно возобновляемая регенерация,что по мнениюКрон-товского имеет принципиальное значение. Кронтовским это обстоятельство было сопоставлено с нек-рыми аналогичными явлениями: повторная ампутация в опытах Чайлда (Chi Id) с планариями Гартмана (Hartmann)—-с амебами и другими простейшими—действовала омолаживающим образом, причем опыты могли быть продолжены неопределенно долгое время без старения индивидуумов. В связи с этим Кронтовским дано объяснение и тому факту, что 18-летняя Каррелевская тканевая культура фибробластов до сих пор растет с прежней активностью, оказывается как бы вечно молодой; на самом деле в течение 2—3-дневиого роста она несколько стареет, но при каждом пассаже искусственно опять возвращается к первоначальному состоянию, что и может повторяться неопределенно долгое время. В пользу такого объяснения говорит и то обстоятельство, что если пассажей с отрезанием периферических частей не производить, то рост культуры по- Рисунок 6. Контуры (слева) и кривая роста чистой культуры эпителия в чашке Карреля. степенно замедляется, клетки превращаются в более зрелые формы, появляются спец. гистологические диференцированные структуры— разного рода волокна и т. п. (Максимов, Фишер, Паркер). Кривая роста «пассажных культур» дана на рисунке 4, потребление сахара—на рисунке 5. Применение метода тканевых культур в различных областях биологии и медицины. Метод тканевых культур с успехом применяется в самых разнообразных областях биологии и медицины. 1. Цитология, гистология, патология. Тенденции современной цитологии—прежде всего изучать объекты по возможности в живом и наименее измененном виде [фиксирование, заключение и т.п. неизбежно вызывает существенные изменения в сложной коллоидной структуре живой протоплазмы (см. Гистологическая техника)]—наилучшим образом отвечает именно метод тканевых культур, поскольку наиболее достоверным доказательством живого (неповрежденного) состояния является рост ткани и размножение клеток (Петерфи). Многими авторами детальнейшим образом изучались структура разных живых клеток в тканевых культурах (при обыкновенном и темнопольном освещении) [см. отдельную таблицу (т. XIV, ст. 375—376), рисунок 6], изменения в цитоплазме и ядре клеток, легко наступающие при всяких неблагоприятных условиях (появление зернистой или иной структуры в ядре, первоначально «оптически пустом», изменение митохондрий и появление различных гранул в цитоплазме и т. п.), при изменении реакции среды (Льюис), осмотического давления (Hogue), при действии лучей Рентгена и радия (Стренджуейс, Канти, Donaldson, Spear) и т. д. В живых активных клетках путем систематической регистрации были обнаружены (Леви, Стренджуейс, Канти и другими) подвижность митохондрий (хондриосом), постоянные изменения их величины и формы (рисунок 7); своеобразные «червеобразные» движения митохондрий прекрасно видны на заме-чательн. кинофильме Канти, демонстрированной Кронтовским на съезде зоологов, анатомов и гистологов в Киеве в 1930 году; изучены различные детали деления клеток при разных условиях и т. д.; на указанной кинофильме особенно хорошо заметен тот стадий митотического деления клетки, к-рый характеризуется как бы «вспузыриванием» («bubbling»), образованием как бы пульсирующих пузырькообразных выступов цитоплазы, имн. др. детали клеточной динамики. Наблюдения за живыми клетками в момент действия на них фиксирующих жидкостей позволили детально изучить, в каких отношениях и насколько изменяется при различной фиксации микроскоп, картина клетки. На отд. таблице (т. XIV, ст. 375—376, рис. 6) видно напр., как «оптически пустое» (даже при темнопольном освещении) ядро клетки, в к-ром заметно лишь ядрышко, при действии смеси Ценкера приобретает своеобразную структуру, становится матово-зернистым [см. отд. таблицу (т. XIV, ст. 375—376), рис. 6 и 7], как изменяется первоначально бесструктурный фон цитоплазмы, в к-рой отчетливо видны в живой клетке капельки жира и нитевидные хондриосомы, как сглаживаются после фиксации общие контуры клетки, периферия к-рой имела в живом состоянии массу тончайших амебоидных, подвижных псевдоподий и т. д. (Стренджуейс и Канти).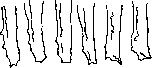
Рисунок 7. Движение и изменение митохондрий в отростке клетки (при темнопольном освещении) в течение 5 мин.
Благодаря методу тканевых культур и «чистым штаммам» отдельные виды клеток получают в наст, время не только морфолог. характеристику (что лежало в основе «классической цитологии»), но и физиологическую, динамическую—по особенностям их движения, скорости роста, по их питанию, реакциям на воздействие и поведению при разных условиях и т. п., что является, по Кар-релю, отличительным признаком «новой цитологии». Исследования над морфологией клеток в тканевых культурах при различных условиях заставили критически отнестись к привычному представлению, что форма клеток есть нечто застывшее, неизменное, характерное для клеток данного вида: достаточно изменения консистенции среды (Уленгут), натяжения (Вейс) и т. п., чтобы типичная эпителиальная клетка превратилась в веретенообразную или отростчатую (Уленгут, Хлопин), сделалась похожей на фибробласт («морфол. конвергенция»); однако она сохраняет при этом свою природу и, помещенная в другие условия, приобретает опять типичную форму эпителия (см. выше опыты Фишера и Иблинга, Дру и др.), в виду чего необходима большая осторожность при суждении (на основании микроскопич. препаратов) о превращении (трансформации) клеток одного вида в другие. В тканевых культурах удается систематически изучать изменения в живых клетках при их ди-ференцировке и проследить совершающиеся при этом коллоидно-химические изменения протоплазмы. В опытах по спермиогенезу in vitro (с культурами из семянников бабочки Samia cecropia) Гольдшмидт, как это показано на рис. 8, наблюдал шаг за шагом развитие in vitro всех стадиев образования спермий и мог экспериментально изучить некоторые факторы спермиогенеза. Как удалось установить, образование осевой нити начинается с того, что на поверхности клетки, обращенной в просвет фоликула, появляется много протоплазматич. выступов, своеобразных псевдоподий (рисунок 8 а—Ъ). Затем один из этих выступов (рисунок 8 Ь) вырастает и спустя некоторое время внезапно превращается в неподвижную нить (рис. 8 с) с утолщением на конце, что обусловлено быстрым изменением коллоидно-хим. структуры, которое Гольдшмидту удавалось вызывать и искусственно. Потом образуется вторая нить (рис. 8 с), и продол-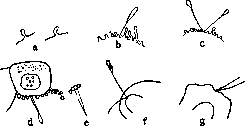
Рисунок 8^ Спермиогенез in vitro.
жается дальнейшее развитие, на течение которого удается оказывать резкое влияние изменением осмотического давления и т. п. (Гольдшмидт). Весьма ценные результаты дал метод тканевых культур и для выяснения проблемы о превращении клеток соединительной ткани и крови, представляющем особый интерес для целого ряда дисциплин: цитологии, гистологии, механики развития, патологии и клин, гематологии. Прослежены изменения гистиоцитов (блуждающих клеток) и превращение их в гипертрофированные, энергично фагоцитирующие полибласты (макрофаги) и превращение в такие же макрофаги моноцитов крови и части лимфоцитов (часть лимфоцитов гибнет), что наблюдается как в культурах из лейкоцитов крови (Авроров и Тимофеевский, Максимов), так и в культурах из лимфоцитов, полученных из ductus thoracicus (Bloom). Сначала гематогенные полибласты (макрофаги) отличаются от полибластов тканевого происхождения (как в эксплянтатах, так и в организме в воспалительных очагах); затем они становятся совершенно сходными (Максимов, Зильбер-берг). В дальнейшем при образовании рубцовой ткани макрофаги и того и другого происхождения превращаются в фибробластоподобные клетки и наконец в настоящие фибробласты (фиброциты), при этом in vitro и in vivo образуются настоящие соединительнотканные (аргирофильные и коллагенные) волокна, как показали Максимов, Блюм и др. Эти наблюдения выяснили и спорный вопрос о происхождении волокон. Зернистые лейкоциты, эмигрирующие из кусочка эксплянтата или эмигрировавшие* из сосудов при воспалении, неспособны к дальнейшему развитию и постепенно погибают. Описан нек-рыми авторами обратный переход фибробластов в гистиоциты (Каррель и Иблинг, Фишер), но это относилось к эмбриональным фибробластам, и тщательному цитологическому анализу эти случаи не подвергались. Незрелые мезенхимные клетки (похожие на фибробласты) могут подвергаться дальнейшим изменениям, фибробласты же представляют собой зрелую форму,неспособную к дальнейшим превращениям (Максимов). Эндотелий, по Максимову, ведет себя аналогично фибробластам; по Льюису,—образует подобно мезепхим-ным клеткам сетчатую ткань. Аналогичные изменения блуждающих клеток—гистиоцитов, моноцитов и лимфоцитов — были установлены в тканевых культурах из разных органов и лейкоцитов крови и при образовании in vitro туб. бугорка (Максимов, Тимофеевский и Беневоленская и др.). Такие опыты производились как с вирулентными туб. бактериями, так и с маловирулентным штаммом BCG-. При этом Льюису и др. удалось проследить непосредственно под микроскопом шаг за шагом многие спорные явления, например образование многоядерных гигантских клеток путем слияния полибластов (рис. 9—зарисовки каждые 10 мин.). Подобное же поведение клеток Тимофеевский описал в тканевых культурах из леп-розных тканей человека, причем он имел возможность наблюдать образование in vitro и типичных «ле-прозных клеток», набитых палочками проказы. Гранулопоэз и эри-тропоэз в тканевых культурах описали Максимов, Беневоленская и др. Систематические исследования упомянутого цикла вопросов дали возможность Максимову создать общее учение о морфологии мезен-химных защитных реакций организма против местных и общих вредностей и дать детальный очерк гистогенеза различных воспалительных реакций. Метод тканевых культур дает возможность изучать основные свойства клеток крови, их проспективные потенции и т. д. при различных формах лейкемий (Авроров, Тимофеевский, Гиршфельд и др.). Менее изучен эпителий разных органов (Шампи, Хлопин) и мезотелий (Мясоедов). По Хлопину, эпителии можно разделить на 3 группы: 1) эпителий желудка, кишечника,-под-желуд. железы, для которого характерны однослойные пласты, 2) эпителий кожи, полости рта и т. д. с тенденцией к образованию многослойных структур, клеточных комплексов типа «жемчужин» и т. п.; 3) разные эпителии мезодермального происхождения (почки, серозные оболочки и т. д.). Исследовано в тканевых культурах развитие гладкомышечных ; волокон (Льюис), нервных волокон (Леви, Olivo, Григорьев и др.), а также реакция и регенерация нервных волокон при повреждениях их иглой микроманипулятора (Леви) и т. п. 2. Физиологические, фармакологические данные. Тщательно изучены скорость и характер амебоидного движения различных клеток животных и человека: лимфоцитов, моноцитов, гистдоцитов и т. д., своеобразное, бы скользящее движение фибробластов, у к-рых, по Каррелю и Иблингу, активным оказывается обычно один конец их веретенообразного тела (характер их передвижения хорошо заметен на кинофильме Канти). У активных моноцитов и блуждающих клеток (гистиоцитов) удалось обнаружить тончайшую, подвижную, ундулирующую мембрану, движение которой заснято и кинематографически (Каррель и Иблинг), похожую на гиалиновую мембрану определенных амебоцитов («хоанолей-коцитов») беспозвоночных (Faure-Fremiet). Особенную ценность представляют данные по анализу мышечного движения. Многими авторами доказано т. н. «миогенное происхождение» ритмич. сокращений сердечной мышцы, т. к. в тканевых культурах из сердца и др. объектах от зародышей и новорожденных животных удалось наблюдать ритмические сокращения отдельных изолированных клеток (Берроуз, М. Льюис, Оливо и др.). Изучены детали сокращений отдельных клеток, их реакция на укол иглой микроманипулятора (М. Льюис), ритмические сокращения эксплянтатов из разных частей сердца (Кон) и т. д. Ритм сокращений обычно довольно правильный. На рис. 10 представлены зарегистрированные Крон-товским (на ленте кимографа) сокращения 2 частей одного эксплянтата из сердца куриного зародыша: а—136 сокращений в 1 мин. при 39°, 6—104. При неблагоприятных условиях вторая часть обнаружила расстройство ритмической деятельности—б иг еминию (рис. 10—Ь). Сокращения продолжаются и при значительном понижении t , напр. при 22° 11—20 сокращений в 1 мин., но уже с неправильным ритмом (рис.10аа). Перистальтика наблюдается в эксплянтатах из кусочков кишечника. В тканевых культурах изучался углеводный обмен (см. выше— химич. динамика). Микрохимически было установлено сильное потребление сахара и образование молочной кислоты, белковый обмен (см. выше), дыхание эксплянтатов (Эрдман) и выделение С02 (de Haan), жировой и липоидный обмены, причем отмечена тенденция разных клеток накоплять в тканевых культурах значительное количество капелек нейтрального жира (Кронтовский и Попев), образование и судьба пигмента (Smith, Luna и др.) и т. п. Изучено непосредственное действие на живые клетки и ткани разных фармакологических веществ: хинина (Парташников), мышьяка (Wilson, Рыбинский), цианистого калия (Olivo), свинца, ртути (Bianchini) и мн. др., причем выяснилась обратимость нек-рых изменений, установлено привыкание ткани к мышьяку (Вильсон) и т. п. Ламберт (Lambert) подверг параллельному изучению действие ряда дезинфицирующих веществ на бактерии и тканевые клетки. Кавецкие производили наблюдения над действием сероводородной воды из источников Мацесты и Сергиевских минеральных вод на рост тканевых культур. 3. Внешние факторы — радиобиология и р е н т г е н о би о л о ги я. При помощи метода тканевых культур было исследовано влияние на живые клетки разных физ. факторов—температуры, высушивания ткани, консистенции среды и т. п.; физ.-хим.—реакции среды (рН), осмотического давления, поверхностного натяжения; химических—разных солей, narcotica, хи- «и Рисунок 10. Ритмические сокращения эксплянтатов из сердца (Ьг — бигеми-ния). нина, мышьяка и т. п.; биол. факторов— экстрактов из разных органов, в том числе из желез внутренней секреции и т. д. Тканевые культуры дают возможность судить о непосредственном прямом действии (в организме же всегда получается суммарный эффект) рентгеновских лучей и радия на клетки и ткани (Amato, Wood, Гасуль, Кронтовский и др.); в частности—изучить действие разных доз [3- и у-лучей радия на митозы (Стренджуейс, Канти, Donaldson, Spear и др.),.на рост и углеводный обмен эксплянтатов (Кронтовский, Коломиец, Лебензон и др.). Посредством соответствующего комбинирования опытов in vivo и in vitro удалось показать участие в реализации эффекта помимо непосредственного воздействия различных вторичных факторов (опыты Кронтов-ского, подтвержденные Шубертом, Стрен-джуейсом и Феллем и др.). Клаег'ом количественно изучено влияние ультрафиолетовых лучей на скорость роста штамма фибро-бластов; прямого стимулирующего эффекта при этом не было обнаружено. 4. Онкологические данные. В отношении изучения проблем онкологии метод тканевых культур дал чрезвычайно ценные, а иногда и решающие результаты. Уже один факт возможности получения в виде чистой культуры in vitro штаммов карциномы мыши (Фишер), саркомы крысы (Каррель), сохраняющих при разных условиях (независимо от организма) свою злокачественность и вирулентность, доказал, что существуют особые злокачественные «раковые», «саркоматозные» клетки, и заставил отбросить ряд прежних гипотез и теорий (в частности положение многих выдающихся патолого-анато-мов, что нет раковой клетки, а есть лишь «злокачественное разрастание эпителия»), позволил подвергнуть раковые клетки детальному и всестороннему изучению при разнообразных, нужных для анализа условиях, и притом прямому, тогда как в организме биология раковых клеток обычно доступна лишь непрямым методам исследования. Посредством тканевых культур удается изолировать отдельные виды клеток, к-рые входят в состав опухоли, и в отдельности изучить их свойства, какие из них обладают злокачественностью и т. п. (Каррель). Метод тканевых культур впервые позволил точно выяснить вопрос о скорости пролиферации раковых и нормальных клеток при одинаковых условиях (в организме условия не равны: рост нормальных тканей б. или м. заторможен нормальными тканевыми корреляциями, злокачественные же клетки не. поддаются этим регулирующим влияниям). Оказалось, что раковые и саркомные культуры растут медленнее нормальных тканей (Каррель, Фишер), что говорит против часто даваемого объяснения, согласно к-рому злокачественным опухолям присуща большая энергия роста(в организме при соответствующих условиях нормальные клетки также могут обнаружить очень быструю пролиферацию—при регенерации, заживлении ран и т. п.). С другой стороны энергично растущие in vitro (так сказать на свободе) нормальные тканевые культуры при имплянтации их в организм сейчас же подпадают под дей- ствие регулирующих факторов, и рост их задерживается (Кронтовский), раковые же культуры, привитые в организм, продолжают и в нем безгранично расти, утратив по-видимому способность подчиняться регулирующим воздействиям нормальных тканевых корреляций. Такого рода исследования дают вполне определенные указания, в каком именно отношении раковые клетки биологически отличаются от нормальных. Тканевые культуры дают также важное указание, что атипическое разрастание с внедрением в подлежащую ткань и т. п., на гист. препарате иногда морфологически совершенно не отличимое от настоящего рака (Максимов), может встречаться в культурах из нормальных тканей и органов, напр. из молочной железы (Максимов), мочеточника и мн. др. Одни морфологические исследования оказываются и здесь недостаточными; необходимы биологическое понимание раковой клетки и тщательное изучение ее физиологических свойств. Если к культурам из опухолей прибавить in vitro кусочки из разных нормальных тканей и органов, клетки опухоли прорастают и уничтожают последние (Фишер, Шаравский). По данным Фишера нормальный эпителий может обнаружить in vitro рост (регенерацию) только при наличии относительно большого конгломерата эпителиальных клеток (см. выше), фибробласты— при несколько меньшем количестве клеток; раковые клетки нуждаются для своего роста лишь в небольшом скоплении клеток, Раусовская саркома (см. Куриные саркомы) способна вырасти из одной единственной клетки. Раковые клетки по мнению Фишера отличаются недолговечностью и меньшей устойчивостью по отношению к разным внешним воздействиям (например к повышению парциального давления кислорода и др.). Рост культур раковых клеток мыши может совершаться in vitro в такой среде, как напр. сыворотка без» эмбрионального экстракта, в к-рой нормальные ткани (эпителий и фибробласты) не растут (Фишер); однако клетки саркомы крысы в этом отношении совершенно сходны с культурой нормальных фибробластов (Каррель), в виду чего необходимы дальнейшие исследования. Основную характерную особенность раковых клеток безгранично расти в организме среди нормальных тканей Фишер объясняет на основании своих опытов тем, что 1) раковые клетки для построения своей протоплазмы могут неограниченно пользоваться (подобно лейкоцитам) сывороткой (плазмой) и 2) что для этой цели они могут применять и вещества приходящих с ними в соприкосновение соседних нормальных клеток. Прививки в организм тканевых культур, обнаруживающих in vitro интенсивный, атипичный рост, как правило не дают положительного результата (Кронтовский и др.), даже если прививаются долго росшие in vitro (с повторной регенерацией) культуры непосредственно в мозг (Колесников). Это говорит в пользу того, что отрыв от материнской ткани и продолжительное автономное разрастание ткани сами по себе недостаточны (вопреки нек-рым теориям) для превращения нормальных клеток в злокачественные. Путем продолжительного выращивания культур из эмбриональной селезенки с прибавлением очень незначительных количеств мышьяка (в опытах Лазера—при культивировании в среде с каменноугольным дегтем) Фишеру удалось после прививки такой культуры курице вызвать развитие настоящей саркомы. Оценка этого опыта тре-•бует осторожности, так как положительный опыт остался единичным несмотря на дальнейшие последующие попытки самого Фишера и других авторов (Карреля и др.). Превращение нормальной клетки в злокачественную повидимому необратимый процесс: ни при каких условиях не удалось превратить культуру опухолевых клеток в нормальные (Каррель, Фишер). Микробиология и учение об иммунитете. Из многочисленных относящихся сюда "исследований здесь приводится лишь несколько разнородных примеров. Относительно изучения при помощи тканевых культур различных фильтрующихся virus'ов, отличающихся вообще особенно интимной связью с клетками организма хозяина, можно указать, что в тканевых культурах (инфицированных in vivo или in vitro) можно обнаружить различные virus'bi: оспенной вакцины, полиомиелита, herpes'а, бешенства, сыпного тифа, везикулезного стоматита лошадей и т. д. и что уже и теперь несмотря на несовершенство техники и незнание требуемых условий нек-рые из них удалось культивировать таким путем в течение долгого времени, тогда как при'помощи обычных бактериол. способов культуры их вообще не получены: virus оспенной вакцины в опытах Паркера и Пая (Nye) с инфицированными in vitro эксплянтатами tes-tis путем пассажей удалось культивировать в течение 132 дней, причем сила virus'а по их расчетам увеличилась (в 11-м пассаже) в 51.000 раз. В пользу размножения virus'а говорит также тот интересный факт, что если приготовить культуру из моноцитов в плазме кролика, содержащей ничтожные следы virus'а, то уже после нескольких дней роста virus может быть легко обнаружен обычными способами (прививкой на роговицу и т. д.). В опытах Карреля и Риверса (Rivers) с эмбриональными культурами в чашках Карреля указанный virus размножается в такой степени, что по мнению названных исследователей с одного куриного зародыша, расчлененного на отдельные тканевые культуры, можно бы -получить примерно столько же оспенной вакцины, сколько можно иметь с целого теленка.—Virus herpes Гааген (Haagen) успешно культивирует в тканевых культурах уже 10 месяцев (50 пассажей). В культурах роговицы кролика, инфицированных оспенным вирусом, можно наблюдать развитие типичных телец Гварньери, а при инфекции вирусом herpes 'а—наличие характерных внутриядерных оксифильных включений, причем и те и другие наблюдаются и в клетках, вновь образовавшихся при пролиферации эпителия in vitro. Вирулентность микроорганизмов в тканевых культурах почти не изучена: по указанию Бехера (Ва-cher) вирулентность ослабленных штаммов пневмококков усиливается, т. ч. по его мнению тканевые культуры могут быть использованы с этой целью вместо прививок животным. По опытам Эрдман с куриной чумой вирулентость ослабевает (необходимы дальнейшие исследования с новой техникой, с обращением внимания на бактерицидность плазмы и т. д.). В тканевых культурах из testis, инфицированных virus'ом herpes'а, Гильдемейстер, Гааген и другие наблюдали ослабление вирулентности после 7-го пассажа; но стоило только ввести небольшое техническое улучшение—производить пассажи каждые 3—4 дня вместо 6,—чтобы в 15—16 пассажей не было замечено никакого ослабления вирулентности (опыты продолжаются дальше). Имеются интересные наблюдения о поведении амеб, разных видов Trichomonas (Hogue) среди тканевых клеток in vitro, бактериофага (Дрезель и М. Льюис) и т. п. «7 Из области изучения иммунитета можно отметить, что в тканевых культурах наблюдалось образование специфических антител: гемолизинов, бактериолизинов, аглюти-нинов, преципитинов. При современной технике на тканевых культурах можно количественно изучить действие последовательно уменьшающихся доз разных токсинов. По различию в скорости роста половинок культур чистого штамма эмбриональных фибро-бластов, из которых одна половина подвергнута действию токсина, легко можно обнаружить влияние дифтерийного токсина в разв едении 1:100.000,1:1 млн. (Кронтовский) и т. п. (при прибавлении к среде). Даже простое прибавление дифтерийного токсина к жидкой части среды над культурами, растущими в чашке Карреля, отчётливо действует на рост (Кронтовский и Яцимир ская-Кр онто-вская); это лучше В всего заметно по кривым роста (рис. 11). Действие токсина можно нейтрализовать антитоксической сывороткой (Левадити). Наметилась возможность и более глубоко проникнуть в механизм действия дифтерийного токсина на жизненные процессы: опыты Кронтовского и Яцимирской-Кронтовской показали ясное действие дифтерийного токсина на химич. динамику тканевых культур, на потребление сахара, образование молочной кислоты (см. табл.). ММ3 1/ / /, -""у. loooo зфт.то Д н и l 2 3 4 Рисунок 11. Влияние дифтерийного токсина на рост культур фибро-бластов. Тканевые культуры в чашках Карреля Поверхность эксплянтированного I куслчка в мм%..........: i,o ! i,o Поверхность всей 3-дневной ткане- . '! вой культуры...........! 8,5 | 1,0 Потребление сахара в ли % .... ' 73,0 ' 49,5 Образование молочной к-ты в мг %. 23,1 ! 10,8 а—тканевые культуры контрольные Ь— » » +0,15% дифтерийн. токсина Опыты с тканевыми культурами являются без сомнения весьма ценным методом и для более точного анализа гуморальных и тканевых факторов иммунитета, как это видно из ряда работ Ламберта, Риверса, Гаагена, Мукенфуса и других по иммунитету при оспе, Кронтовского 11 Гаха—при сыпном тифе и мн. др. Лит.: Монографии и обзоры.— К р о н т о в -с к и й А., Обзор работ по тканевым культурам, Центр, мед. гк., т. IV, в. 2, 1929 (обширная лит.); он ж е, Химическая динамика регенерирующих и бла-стоматозных тканей, Врач, дело, 1929, № 20; Кронтовский А. и Полев Л., Метод тканевых культур, ч. 1—2, Киев, 1917 (лит.); Максимов А., О культивировании in vitro соединительной ткани взрослых млекопитающих, Рус. архив анат., гист. и эмбриол., том I, 1916; Румянцев А., Тканевые культуры и их значение в биологии, М.—Л., 1930; Тимофеевский А., Значение тканевых культур для решения нек-рых проблем гематологии, Сиб. арх. теор. и клин, мед., т.1, кн. 3—4, 1926; В i s с е g - Б. М. Э. т. XV. Не V. und Juhasz-Schaffer A., Die Ge-webezuchtung in vitro, В., 1928; Carrel A., Tissue culture and cell physiology, Physiol. reviews, v. IV, 1924; Faure-Fremiet E.etEphrussi В., La culture des tissus «in vitro», Ann. d'anat. pathol., t. V, 1928; Fischer A., Gewebeziichtung, Munchen, 1930 (лит.); Krontowsky A., Explantation u. deren Ergebnisse f. d. normale u. pathol. Physiologie, Erg. d. Physiol., B. XXVI, 1928 (лит.); Levi G., Vita autonoma di parti dell* organismo—La coltiva-zione dei tessuti, Bologna, 1922; Lewis W. a. Lewis M., Behavior of cells in tissue cultures (General cytology, ed. by E. Cowdry, Chicago, 1924, лит.). Методика.—Carrel A., Tissue cultures in the study of viruses (Filterable viruses, ed. by Th. Rivers, London, 1928); D e m u t h Fr., Praktikum der Ziich-tung von Warmblutergewebe in vitro, Munchen, 1929; Erdmann R., Praktikum der Gewebepflege oder Explantation besonders der Gewebeziichtung, В., 1922; Fischer A., Technik der Gewebeziichtung (Hndb. d. biol. Arbeitsmethoden, hrsg. v. E. Abderhalden, Abt. 5, T. 1, B.—Wien, 1927); о н ж е, Technik uiid Methodik der Gewebskulturen (Hndb. d. pathogenen Mikroorganismen, hrsg. v. W. Kolle, R. Kraus u. P. Uhlenhuth, B. X, 1929); Levi G., Gewebeziichtuii'? (Methodik der wissenschaftlichen Biologie, hrsg. v. T. Peterfi, B. I, B., 1928); Strangeways Т., Tissue culture in relation to growth and differentiation, L., 1924. Периодическое издание. —- Archiv fur experimented Zellforschung besonders Gewebeziichtung, Jena, С 1925. А. Кронтовский.
Большая медицинская энциклопедия. 1970.
Культуры тканей — эксплантация (биол.), метод длительного сохранения в живом состоянии клеток, тканей, небольших органов или их частей, выделенных из организма человека, животных или растений. Первые успешные опыты по К. т. осуществил в 1907 американский… … Большая советская энциклопедия
Микрорастение плодовой (ягодной) культуры — Микрорастение плодовой [ягодной] культуры: растение плодовой [ягодной] культуры, полученное путем укоренения побегов, выращенных с использованием метода культуры тканей... Источник: МАТЕРИАЛ ПЛОДОВЫХ И ЯГОДНЫХ КУЛЬТУР ПОСАДОЧНЫЙ. ТЕРМИНЫ И… … Официальная терминология
микрорастение плодовой [ягодной] культуры — Растение плодовой [ягодной] культуры, полученное путем укоренения побегов, выращенных с использованием метода культуры тканей. [ГОСТ Р 53044 2008] Тематики посадочный материал плодовых и ягодных культур … Справочник технического переводчика
КУЛЬТУРА ТКАНЕЙ — Рис. 1. Культуры ткани почек свиньи и крупного рогатого скота. Рис. 1. Культуры ткани почек свиньи (а) и крупного рогатого скота (б). Три морфологически различных типа клеток. культура тканей, тканевая культура, метод выращивания вне организма в… … Ветеринарный энциклопедический словарь
КУЛЬТУРА КЛЕТОК И ТКАНЕЙ — У растений искусств. индуцирование деления клеток в изолированных кусочках (локусах) растит, тканей или целых органах р ний (пыльниках, семяпочках и др.). Начало изучению метода К. к. и т. положено трудами Ф. Уайта и Р. Готре (нач. 20 в.), затем… … Сельско-хозяйственный энциклопедический словарь
культура клеток и тканей — культура клеток и тканей. У растений — искусственное индуцирование деления клеток в изолированных кусочках (локусах) растительных тканей или целых органах растений (пыльниках, семяпочках и др.). Начало изучению метода К. к. и т. положено… … Сельское хозяйство. Большой энциклопедический словарь
культура клеток и тканей — метод сохранения в жизнеспособном состоянии клеток, участков тканей, органов или их частей вне организма. Во 2 й пол. 19 в. развитие микробиологии, прежде всего медицинской (необходимость выделения и изучения микробов, вызывающих инфекционные… … Биологический энциклопедический словарь
КУЛЬТУРА ТКАНЕЙ — эксплантация, метод сохранения жизнеспособности органов или их частей, участков тканей и отд. клеток вне организма. К. т. основана на создании асептич. условий, обеспечивающих питание, газообмен и удаление продуктов обмена культивируемых объектов … Биологический энциклопедический словарь
Доколумбовы культуры — Скульптурная трубки доколумбовой эпохи, найденная археологами у кургана Накучи, штат Джорджия, США Одна из пирамид на верхнем уровне руин Йашчилана, Мексика Гигантские скульптур … Википедия
Доколумбовы цивилизации и культуры — Скульптурная трубки доколумбовой эпохи, найденная археологами у кургана Накучи, штат Джорджия, США Одна из пирамид на верхнем уровне руин Йашчилана, Мексика Гигантские скульптур … Википедия
КУЛЬТУРЫ ТКАНЕЙ
18+
© Академик, 2000-2025
- Обратная связь: Техподдержка, Реклама на сайте
Экспорт словарей на сайты, сделанные на PHP, Joomla, Drupal, WordPress, MODx.